Before a cell can divide, it must first replicate its genome, or the sum total of its DNA, so that each of the two daughter cells inherits its own copy. In 1953, James Watson and Francis Crick completed their landmark paper on the DNA double helix with what would become a famous prediction about genome replication:
It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material.
Watson and Crick conjectured that the two strands of the parent DNA molecule unwind during replication, and then each “parent” strand acts as a template strand for the synthesis of a new complementary strand of “daughter” DNA (see figure below). Their hypothesis is an interesting one, but is it true?
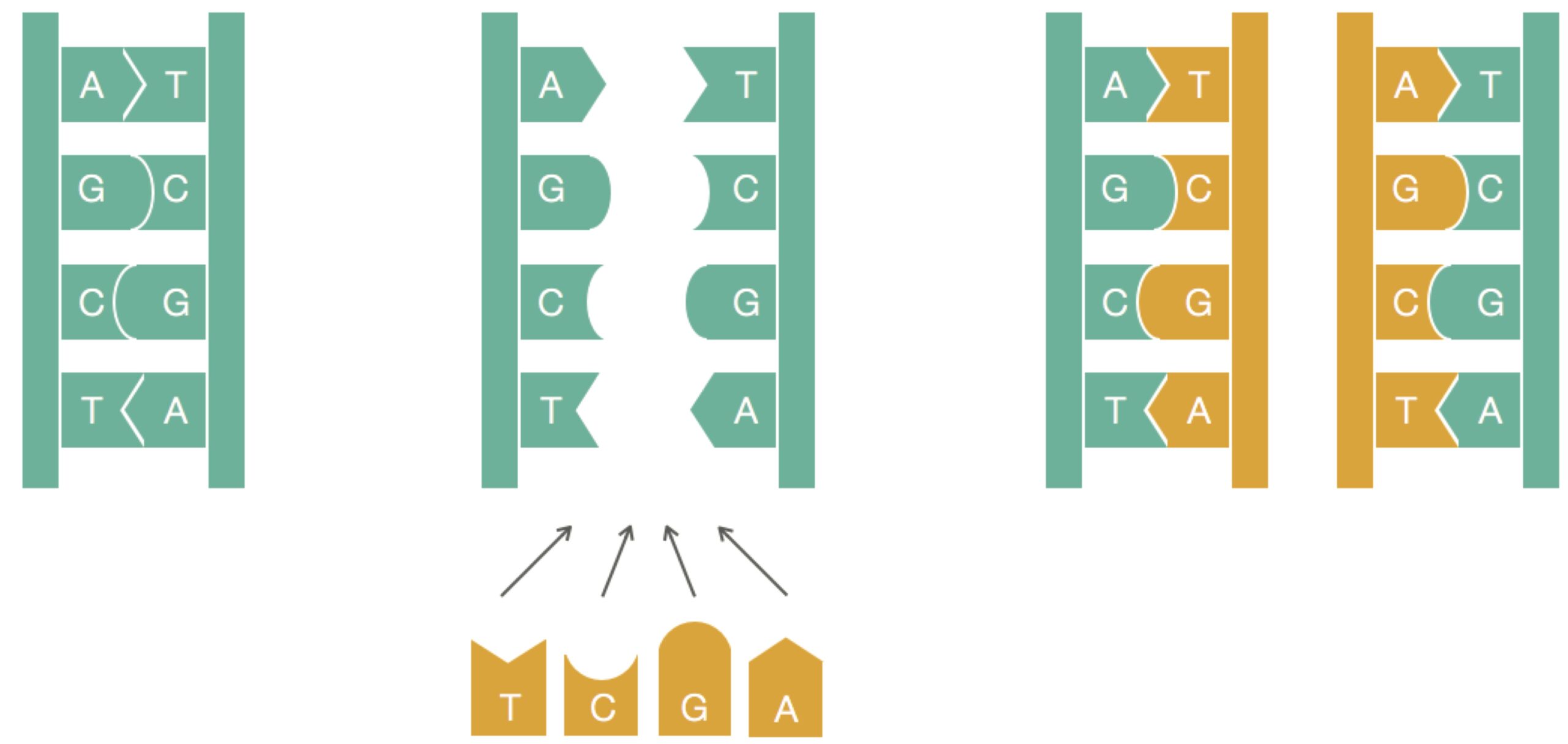
A) and thymine (T) are complements of each other, as are cytosine (C) and guanine (G). Complementary nucleotides on opposing strands of DNA bind to each other.Three hypothetical models of DNA replication
During the 1950s, scientists debated three proposed models for DNA replication, illustrated in the figure below. The semiconservative hypothesis owed to Watson and Crick’s suggestion that each parent strand acts as its own template for the synthesis of a complementary strand. The conservative hypothesis proposed that the entire double-stranded parent DNA molecule serves as a template for the synthesis of a new DNA molecule, resulting in one molecule with two parent strands and another with two daughter strands. The dispersive hypothesis proposed that some mechanism breaks the DNA into fragments and then splices the fragments together, mixing parent and daughter fragments, so that each of the four resulting strands is a patchwork of parent and daughter DNA.
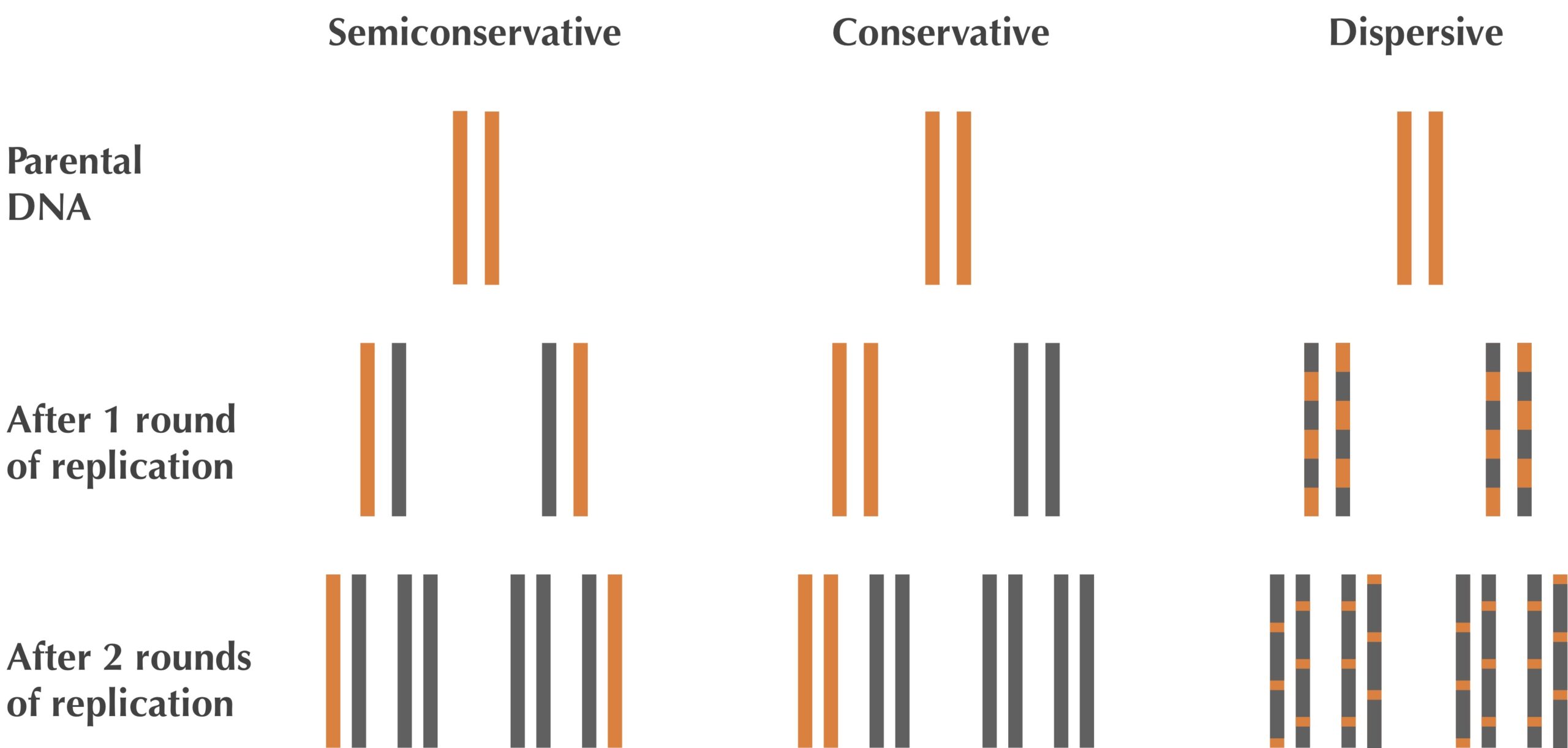
The Meselson-Stahl experiment
The debate would eventually be resolved five years later by Matthew Meselson and Franklin Stahl, who carried out what has become known as “the most beautiful experiment in biology”. Their insight was that one isotope of nitrogen, Nitrogen-14 (14N), is lighter and more abundant than Nitrogen-15 (15N). Knowing that DNA naturally contains 14N, Meselson and Stahl grew E. coli for many rounds of replication in a 15N medium, which caused the bacteria to gain weight as they absorbed the heavier isotope into their DNA. When they were confident that the bacterial DNA was saturated with 15N, they transferred the heavy E. coli cells to a less dense 14N medium.
STOP: What do you think happened when the “heavy” E. coli replicated in the “light” 14N medium for one generation?
After one round of replication, the three hypotheses agreed that half of the nitrogen in the DNA would be the 14N isotope, but they disagreed on how this 14N would be distributed over the DNA molecules. The conservative model predicted that half of the double-stranded molecules of DNA would still have only 15N and therefore be heavier, whereas the other half would have only 14N and be lighter. Yet when they used a centrifuge to separate the E. coli DNA according to weight after one round of replication, all of the DNA had the same density! Meselson and Stahl had just refuted the conservative hypothesis.
STOP: Note that both the dispersive and semiconservative hypotheses predicted that all of the DNA after one round of replication would have the same density. What would these models predict about the density of E. coli DNA after two rounds of replication?
The dispersive model predicts that after two replication cycles, every strand of DNA should contain about 25% ¹⁵N and about 75% ¹⁴N. In other words, all the DNA should still have the same density.
And yet when Meselson and Stahl spun the centrifuge after two rounds of E. coli replication, they observed two different densities of DNA. This is exactly what the semiconservative model predicted. After one cycle, every cell should possess one 14N strand and one 15N strand; after two cycles, half of the DNA molecules should have one 14N strand and one 15N strand, while the other half should have two 14N strands, producing the two different densities that they noticed.
STOP: What does the semi-conservative model predict about the density of E. coli DNA after three rounds of replication?
Meselson and Stahl had refuted the conservative and dispersive hypotheses of replication, and yet they wanted to make sure that the semiconservative hypothesis was confirmed by further E. coli replication. This model predicted that after three rounds of replication, one-quarter of the DNA molecules should still have a strand consisting of 15N, so that 25% of the DNA would be heavier, whereas the remaining 75% would have only 14N and be lighter. This is indeed what Meselson and Stahl witnessed in the lab.
Where is the computation?
We must point out that Meselson and Stahl’s experiment does not offer the concrete finality of the mathematical proofs that we saw in Chapter 0 with the work of the Ancient Greeks. After all, they only established the semiconservative hypothesis for one organism of E. coli; this hypothesis may be violated for other E. coli organisms, or other bacteria, or in turn to eukaryotic organisms like you and me. Nevertheless, the semiconservative hypothesis has been witnessed in all living things since their beautiful experiment.
And yet, you are probably wondering where the computation is hiding. How can we find anything interesting with respect to programming within the replication of DNA? Read on to find out, or take a short detour to watch a lovely interview with Meselson and Stahl, recorded over 60 years after their experiment.

